Humans are part of the wider mammalian group called Primates, and more narrowly are most closely related to the living great apes, in particular the chimpanzees of tropical Africa. This close relationship can be confirmed both from anatomy and from the DNA in our genomes, which can be used to estimate that our ancestors diverged from those of chimpanzees between about 5 and 9 million years ago. Although fossil evidence from that time period is rare, remains from Chad, Kenya and Ethiopia indicate that creatures living there already displayed the human features of small canine teeth and a propensity to walk upright on two legs more than four million years ago. By 3 million years ago there is clear evidence of the full modification of the skeleton for habitual bipedalism in a number of species of australopithecines ("Southern apes"), as well as the earliest signs of stone tool production, and by 2.5 million years the first fossils assignable to our genus Homo are present in East Africa, along with unequivocal evidence of the processing of animal carcasses for meat, using stone tools.
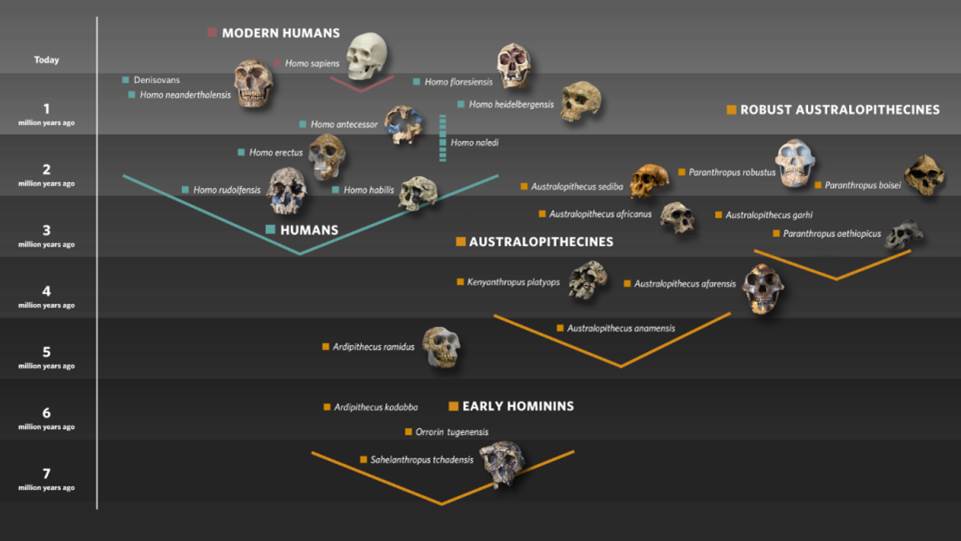
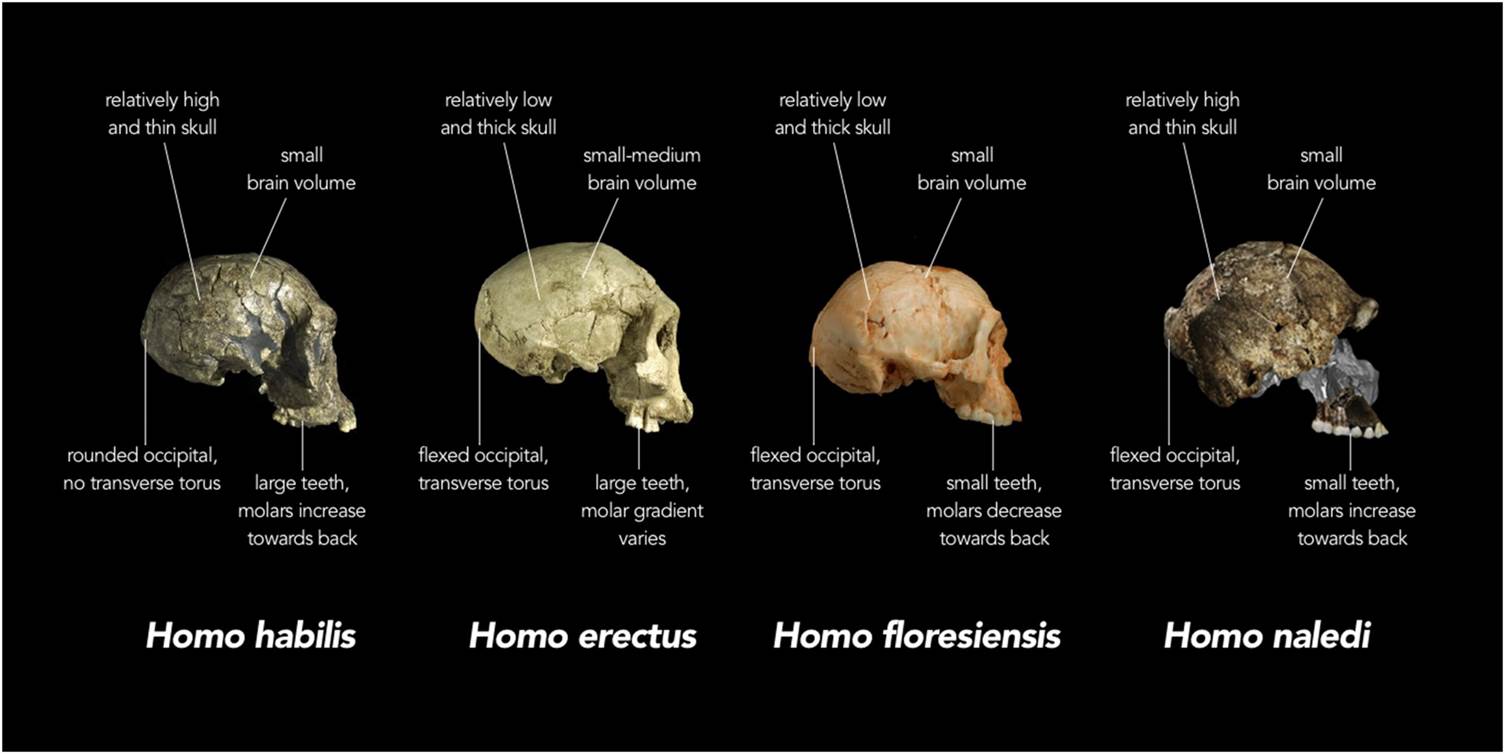
The term "Human" can be defined in many ways, but with respect to what will be preserved in the fossil record, we can look for such things as a large brain in relation to body size, a reduction in the size of the teeth, jaws and associated musculature, and a skeleton characterised by a human-shaped rib cage, the loss of climbing adaptations in the upper body, relatively long legs, and an extended period of childhood growth. The first fossils which are complete enough to display most of this pattern are from East Africa and western Asia, dating from 1.5 to 1.9 million years, and these are generally assigned to the species Homo erectus. This was the first really widespread human species, and the name was originally given to fossils found far away from Africa, on the Indonesian island of Java. Homo erectus is believed to have evolved in Africa about 2 million years ago, and soon after that it dispersed to western Asia (as represented by fossils at the Georgian site of Dmanisi), East Asia (e.g. China) and south east Asia (e.g. Java). Those dispersals were probably facilitated by environmental changes which gave early humans feeding opportunities over wider distances, and it is likely that the spread across what is now island south east Asia as far as Java was possible because of land bridges existing at that time.
Homo erectus individuals were already comparable to humans of today in body size and shape, but with much wider hips and greater muscularity, thicker skulls characterised by smaller brains, larger faces capped by a strong brow ridge over the eye sockets, and chinless lower jaws. Technologically, early Homo erectus was equipped with a simple stone tool kit which looks better suited to scavenging, but later erectus peoples produced a wider range of stone tools and may well have been active hunters. Some experts believe that the more linear and long-legged physique of Homo erectus evolved to facilitate long-distance movements across open country to acquire meat. Nevertheless, it is likely that plant resources have also always been important to humans throughout their evolution.

As mentioned above, Homo erectus is well-known from fossils, but until recently other putative early human species such as Homo habilis and Homo rudolfensis were less well represented in the record, meaning that their place in human evolution remains uncertain. However, in the last couple of years a wealth of material representing a new and primitive human species, Homo naledi, has been recovered from deep within the Rising Star cave system in South Africa. How these remains arrived in a virtually inaccessible chamber is unknown, although there are claims that they were somehow intentionally placed there. There are currently over 1500 fossils representing at least 15 individuals, both adults and children, and this species seems more primitive in several ways than Homo erectus, with an ape-sized brain and signs that the upper body and curved fingers were still adapted to life in the trees. Yet the hips, legs and feet demonstrate a clear bodily commitment to walking regularly on two legs, while the teeth and jaws have Homo-like shapes and proportions, suggesting a human-like diet. Unfortunately, the naledi material is currently undated so it is difficult to place this newly-discovered species in the big picture of human evolution, yet the primitive aspects of its morphology suggest it should be quite basal on the human evolutionary tree, perhaps close to an ancestral form for Homo erectus .
Homo erectus was long-lived and successful, and gave rise to various descendant species in the Old World. One example may be the controversial form nicknamed "The Hobbit", Homo floresiensis, so far known solely from the island of Flores, about 500 km to the East of Java. This species was named in 2004 from remains which included a ~60,000 year old diminutive partial skeleton with skull and lower jaw, excavated from a deep pit in Liang Bua cave. It has been argued that this small-bodied and small-brained species is actually a descendant of Homo erectus, which was then subjected to a process called insular dwarfing, when it arrived by some means on the remote island of Flores. This view has been reinforced by the discovery of similarly small jaw and dental remains at the ~700,000 year old site of Mata Menge in central Flores. Yet in reality, the origin and fate of Homo floresiensis are still clouded in mystery .
In Europe it seems likely that Homo erectus gave rise to the species Homo antecessor, currently only known from the site of Gran Dolina, in the Atapuerca hills of northern Spain. The material is dated to about 850,000 years ago, and consists of more than 150 fossils, predominantly from immature individuals. Most of the bones display butchery marks consistent with them having been processed to extract meat and marrow, although the exact circumstances of this apparent cannibalism are, of course, unknown. The mainly Spanish team who named and studied the material has suggested antecessor could have been the last common ancestor of the subsequent Neanderthal and Homo sapiens lineages, with the flat face and hollowed cheekbones of several individuals presaging the morphology found in modern humans. I will return to this issue of ancestry and descent later.
A subsequent species, named from a fossil jawbone found in Germany in 1907, is Homo heidelbergensis. In the view of some scientists, including me, this species was widespread in Eurasia and Africa from about 600-300,000 years ago, with representative remains including fossils from Germany (Mauer, near Heidelberg), England (Boxgrove), France (Arago), Italy (Ceprano), Greece (Petralona), Ethiopia (Bodo), Zambia (Broken Hill) and South Africa (Elandsfontein). It may also have existed in Asia, as indicated by fossils such as Narmada (India) and Yunxian (China), but this is still unclear. Homo heidelbergensis was typified by a very large body size in males and a brain size overlapping that of humans today, while the body shape was human-like, but with a very broad pelvis and shoulders. The skull was large but long and low, with large brow ridges and a chinless lower jaw. I have argued for many years that heidelbergensis rather than antecessor was the probable last common ancestor of Neanderthals and modern humans, with the divergence of the respective descendant lineages in Eurasia and Africa developing after about 500,000 years ago. However, new fossil and genetic data, described next, are making me reconsider this view.
The hills of Atapuerca are not only famous for the Gran Dolina site which produced the Homo antecessor fossils. Deep within them is the Cueva Mayor cave system and a chamber called, with good reason, the Sima de los Huesos ("Pit of the Bones"). Since 1976 a large number of cave bear and human fossils have been recovered from the Sima, with the human material now numbered at over 6,500 fossils, representing about 28 individuals. The human remains consist of jumbled partial or nearly complete skeletons, mainly those of adolescents and young adults, and were previously claimed to be about 600,000 years old, and to represent Homo heidelbergensis. However, they display clear affinities to subsequent Neanderthals in details of the skull, face, jaws and especially their teeth and now are dated to about 400,000 years instead. Moreover, remarkably, these remains have yielded ancient DNA which places them firmly on the Neanderthal genetic lineage, in line with their morphology, and their status as very early Neanderthals has led me to think that Homo heidelbergensis might be too close in age to the Sima people to represent an ancestor, when the descendant is already well along the Neanderthal lineage. Additionally, new genetic calibrations, partly based on the Sima data, now place the Neanderthal-sapiens divergence at about 650,000 years ago, closer to the time of antecessor. So I am currently unsure whether the last common ancestor of the later species was more like Homo heidelbergensis or Homo antecessor, and I am also unsure on which continent that ultimate ancestor lived.
Thus the Neanderthal lineage was already well-established in Europe by 400,000 years ago, and we can track the evolution of this group through early fossils like Swanscombe (England) and Steinheim (Germany), through material like Ehringsdorf (Germany) and Biache (France), to the best known examples from Europe and western Asia in the period 40-130,000 years ago. The late Neanderthals retained the wide bodies of heidelbergensis but were shorter in stature, with limb proportions which minimised surface area, presumably to conserve heat under the predominantly colder conditions of the last 200,000 years. Their skulls were still long and low with a definite brow ridge, but their brains were larger than the modern average, although not larger when body size is taken into account. The Neanderthal face was dominated by a wide and projecting nose, while the front teeth were somewhat enlarged, perhaps because they were regularly used like a third hand in the processing of food and other materials. The teeth were placed quite far forwards in the jaws, while the lower jaw generally lacked any sign of a projecting chin. Ancient DNA began to be recovered from Neanderthal fossils in 1997, and this has led on to the reconstruction of several complete genomes in the last few years . These indicate that Neanderthals ranging from Spain to Siberia were low in numbers and diversity during the last 20,000 years before they went physically extinct about 39,000 years ago. This may be because regular and extreme climate fluctuations during the last 100,000 years continually fragmented Neanderthal populations, but the reasons for their eventual extinction are still unclear (I will discuss this further below).
While the Neanderthals were evolving in Eurasia, the lineage of Homo sapiens was evolving in Africa. Early examples of this evolution may be represented by fossils like Eliye Springs (Kenya), Florisbad (South Africa) and Jebel Irhoud (Morocco), and by 160-195,000 years ago there were humans close to the modern anatomical pattern in Ethiopia, as represented by fossils from Omo Kibish and Herto. This modern pattern is typified by a more linear body build, with narrower hips and shoulders, and a braincase which is rather globular in shape and lacking a strong brow ridge. The face is smaller and retracted under the skull, and there is a chin on the lower jaw, even in infants. By 100,000 years ago humans in Africa were a very diverse group, and some emerged into western Asia, as represented by fossils from the Israeli sites of Skhul and Qafzeh. These groups may have spread further than the Middle East but it is generally acknowledged from genetic data that the main dispersal of Homo sapiens from Africa only took place about 60,000 years ago. By 42,000 years ago modern humans were penetrating the territories of the last Neanderthals in Europe, and on the other side of the world they had used boats to reach the remote regions of New Guinea and Australia (the colonisation of the Americas only began in the last 20,000 years, however). So what facilitated the successful spread of modern humans, and what happened to the Neanderthals and other descendants of Homo erectus outside of Africa?
I mentioned Homo floresiensis earlier, as an enigmatic form so far found only on the island of Flores. We know that Homo erectus was a long term inhabitant of the island of Java, with fossils dating from about 1.5 million years down to perhaps 250,000 years ago, while the species may have been present in China until at least 400,000 years ago. But its date of disappearance in Indonesia and China is currently unknown, and there are fragmentary and more recent remains that might represent the species from places like Taiwan and The Philippines, as well as stone tools perhaps 150,000 years old on the island of Sulawesi. Additionally, much further north in Denisova Cave in Siberia, fragmentary fossils including isolated large teeth were tested for ancient DNA in 2010, revealing a new kind of human colloquially known as "Denisovans" . Their DNA identifies them as a distant relative of the Neanderthals, perhaps diverging from their lineage about 450,000 years ago, but we know little about them beyond their excellently-preserved genomes. There are also fossils in China from the period between 60,000 and 350,000 years ago such as the partial skulls from Dali, Maba and Xujiayao, and the skull and large-bodied skeleton of a woman from Jinniushan, but these are difficult to classify, in some ways resembling Homo erectus, in other ways antecessor, heidelbergensis or the Neanderthals. Perhaps some of these are Denisovans, but without their DNA, we cannot tell.
Many scientists like me considered that the spread of modern humans from Africa about 60,000 years ago had led to the almost total replacement of the other human groups on Earth. While I regard forms like the Neanderthals as distinct species from us, I never considered that separation was wide enough to prevent some level of interbreeding, but I thought it unlikely that a small level of hybridisation from more than 40,000 years ago could have survived to the present day. However, I was wrong about that! In the last few years we have learnt that humans outside of Africa carry around 2% Neanderthal-derived DNA in their genomes, from interbreeding events about 50,000 years ago. In addition, some populations in Oceania (especially Papua-New Guinea and Australia) carry an even higher level of DNA in their genomes from ancient interbreeding with a Denisovan-like population. Since the ancestors of Oceanians are thought to have travelled through southern and then south east Asia rather than northern Asia, this indicates that there was probably a Denisovan-like population living in south east Asia as well as Siberia, where the interbreeding occurred. There has also been speculation that sub-Saharan African genomes carry evidence of introgression from an archaic human species such as Homo heidelbergensis. Moreover, recent genome analysis of Andamanese Islanders and other groups has led researchers to propose that extant populations in south east Asia carry evidence of yet another interbreeding event, with a more ancient population than Neanderthals or Denisovans. While there are several possible sources for this interbreeding including Homo floresiensis, speculation is centring on Homo erectus as the most likely candidate. If confirmed, that would provide definite evidence of the late survival of this species. While most of the introgressed DNA in our genomes seems to be non-functional, some elements may have had health benefits in the past in terms of disease-resistance and faster blood-clotting, while there may be negative effects today in terms of some auto-immune conditions.
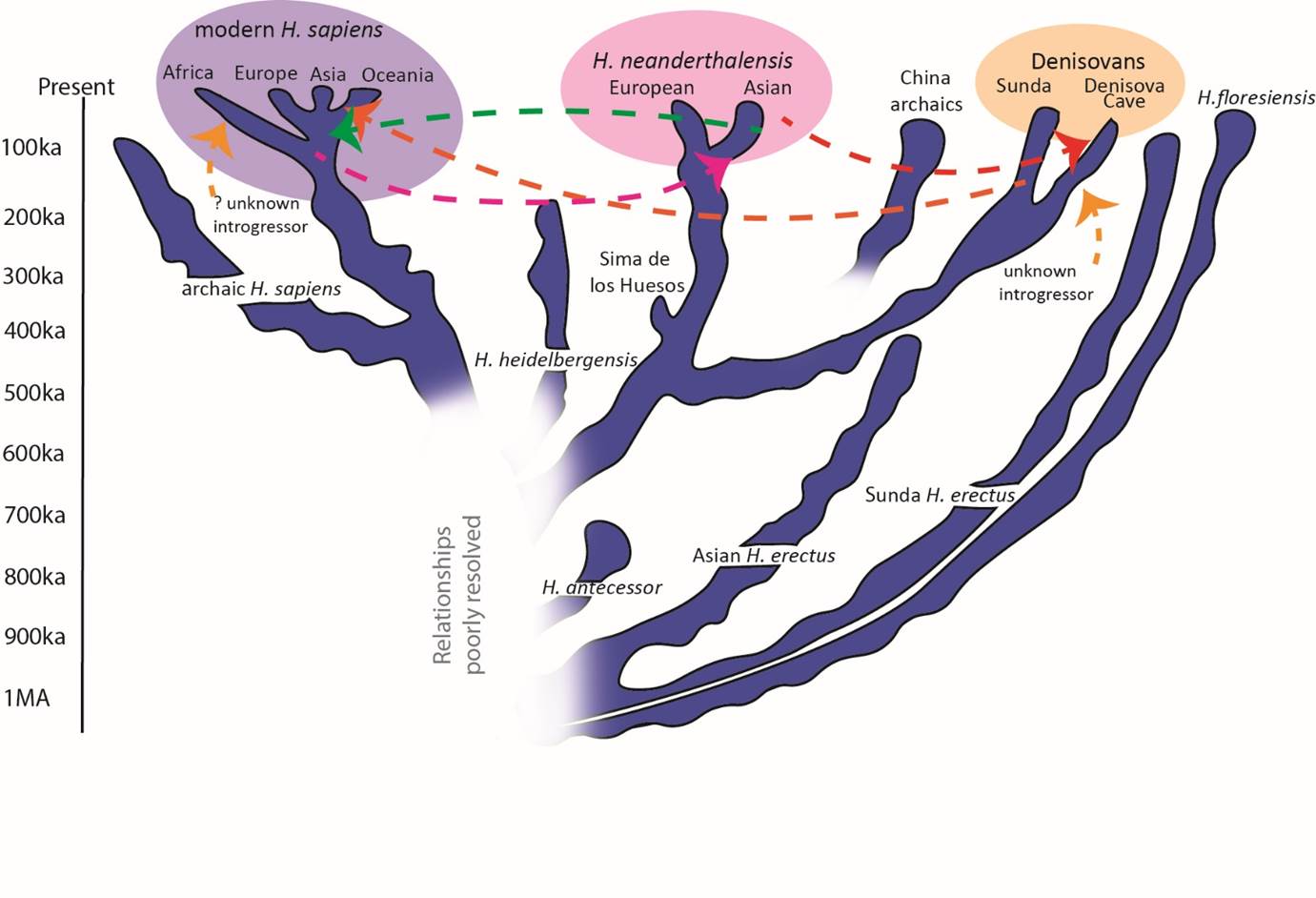
Finally, we need to consider why Homo sapiens is the only surviving species out of the many discussed above . One possibility is that modern humans simply out-competed the other species where they overlapped in time and space, through more effective hunting and gathering, and the occupation of the most favourable environments. Moreover, the genetic data show that at least some of the other human populations were low in numbers and diversity at the time of the modern human dispersal from Africa, and thus these populations were perhaps especially vulnerable to extinction. Yet although the replacement of peoples like the Neanderthals and Denisovans may have happened relatively quickly, it was not an overnight event, and genetic data show that it was certainly not a total replacement in all the regions outside of Africa. To better understand the reasons for our eventual success we are not only going to require much more genetic data from the fossils, but also a far richer fossil and behavioural record from regions like Asia and south east Asia, as well as Africa.
Further reading on this topic:
- Foley R, Martin L, Lahr M, and Stringer C (2016) Major transitions in human evolution Phil. Trans. R. Soc. B 371: 20150229
- Stringer C (2016) The origin and evolution of Homo sapiens Phil. Trans. R. Soc. B 371: 20150237
Other Articles by This Author
Further Articles in  Paleobiology
Paleobiology
 This work is distributed under a Creative Commons Attribution ShareAlike 4.0 International License (CC BY-SA 4.0). Its use, distribution and reproduction in other forums is permitted provided both the original author(s) and Capeia are accredited, and the license is unchanged (CC BY-SA 4.0). Images or other third party material in this article are covered under the article’s Creative Commons license unless otherwise indicated; if the material is not covered under a Creative Commons license, users will need to obtain permission from the license holder before it can be reproduced.
This work is distributed under a Creative Commons Attribution ShareAlike 4.0 International License (CC BY-SA 4.0). Its use, distribution and reproduction in other forums is permitted provided both the original author(s) and Capeia are accredited, and the license is unchanged (CC BY-SA 4.0). Images or other third party material in this article are covered under the article’s Creative Commons license unless otherwise indicated; if the material is not covered under a Creative Commons license, users will need to obtain permission from the license holder before it can be reproduced.
Responses