Braided Streams: Evolutionary Dynamics Among Pleistocene Hominins in East Asia
By Sheela Athreya in Paleobiology |
Paleobiology | Where Did Humans Evolve?
You’ve probably heard it said many times: Africa is the cradle of humankind. For the most part, this is true. Our immediate hominin ancestors evolved in Africa, and so too did our own genus, Homo. But what about our species, Homo sapiens?
In the early 1900s, most scientists actually thought that Asia was the original homeland of Homo sapiens. There are several historical reasons why the focus slowly shifted to Africa but for the past 30 years, the origin of "modern" human biology and behavior has solidly been seen as originating there. Most paleoanthropologists now believe that our species only started occupying Eurasia following dispersals out of Africa in the Late Pleistocene (128,000-12,000 years before present/b.p.). The archaic populations already living there, such as Homo erectus, Homo heidelbergensis, and Neanderthals are believed to have gone extinct when Homo sapiens spread throughout the globe (replacement scenario). And in this scenario, East Asia is viewed as an "evolutionary backwater" in terms of human evolution, while Africa and Europe are at center stage.
The fossil records from China and Indonesia are very rich and have been known since the late 1800s. They tell a different story than the one proposed by the above so-called "Out of Africa" model. Most students of anthropology hear about "Multiregional Evolution" as an alternative to Out of Africa . It’s often misunderstood as proposing parallel evolution in different regions for local Homo sapiens populations through time. For this reason, it’s also often dismissed because how likely is it that "Asians" evolved separately but at the same rate and in the same direction as "Europeans" and "Africans" to lead to the unified human species today? But this is not what the idea of multiregionalism is actually proposing . It is actually founded on the idea of gene flow among these populations – something that prevents two groups from splitting off and becoming separate species. So in reality the idea of multiregional evolution is a complex scenario involving some continuity through time in the genetic contribution of ancient populations in each region to the evolution of Homo sapiens in that region today, but it also very much expects that the evolutionary processes of natural selection, genetic drift, and especially gene flow shaped these populations. When understood that way, it makes sense that there was continuity between the earlier and later populations within each region. That’s why the model is also known as Regional Continuity model: no regional population evolved in isolation, nor did any face complete extinction and replacement by dispersing Africans.
Looking Beyond Africa
Homo sapiens evolution took place during a time period known as the Pleistocene, or Ice Age. It spans from 2.58 million years b.p. to 12,000 years b.p.. The Middle and Late Pleistocene fossil record from China is vast – perhaps as vast as the African record for this time period. The dry, cold climate that China periodically experienced during the Pleistocene, along with its numerous limestone caves, seems to have provided the perfect combination of suitable habitation sites for our hominin ancestors, and excellent preservation for today’s fossil hunting paleoanthropologists.
The best-known hominin fossil locality in China is the home of "Peking Man". For generations, the inhabitants of the village of Zhoukoudian (about 41 km southwest of Beijing) knew about the fossil bones in nearby caves. Beginning in the early 1920s, European paleontologists searching for evidence of human evolution homed in on these caves, and from 1921 until the mid 1940s the site was meticulously excavated by a collaborative Chinese-European team. The original fossils were unfortunately lost during WWII when the Japanese invaded China; only high quality first-generation casts remain . Recent advanced dating techniques confirm that the oldest levels at Zhoukoudian contain Homo erectus fossils and date as far back as approximately 700,000 years while the youngest fossils from the site are Homo sapiens and date to around 35,000 years ago. Only these two hominin populations (or species, depending on your point of view) were found. Across this nearly 670,000 year span we see important changes in the biology of these populations, but also some continuity of traits that are characteristic of all Asian populations. Although this is just one site, it is so rich in paleoanthropological material and so well studied that it serves as a baseline for more recent studies of human evolution in East Asia.
In the past ten years several new sites have added considerably to the picture first painted by Zhoukoudian. These, together with sites as recent as 11,000 years b.p. reveal that populations that lived in China throughout the last glacial period had anatomical traits that in some cases align them with living Chinese but in some cases do not resemble any other contemporaneous population in Eurasia. These fossils point to a more complex picture of human evolution in eastern Eurasia than the simple "Out of Africa vs. Multiregional Evolution" models provided. You could say that the morphology of these populations is "unexpected", but is it really? That would only be true if 30-year-old models defined our expectations, and we had strictly European notions of what early Homo sapiens should look like. In reality these data give us a much clearer, globally inclusive picture of what constitutes “modern humans.”
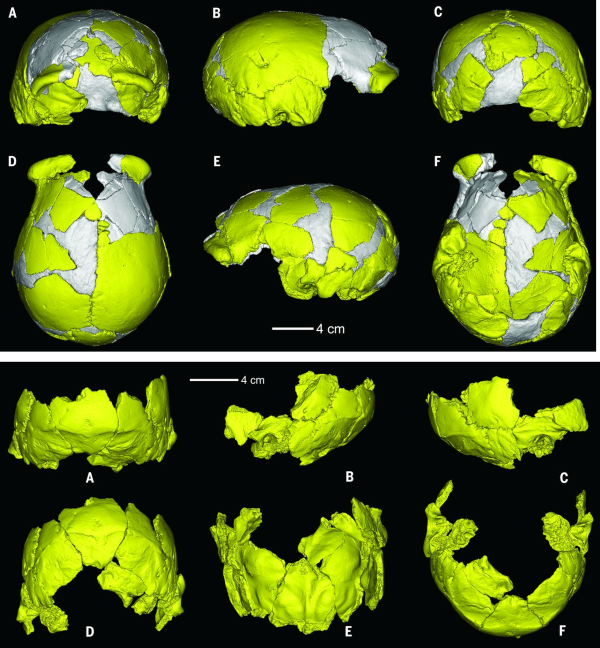
The Earliest Homo sapiens in China: Xuchang
The open-air site of Xuchang in Henan province, central China is a former spring located just south of the Yangtze River. In 2007 two crania were found in archaeological levels directly dated to between 105-125,000 years b.p. . The most complete specimen, Xuchang 1 has a cranial capacity (i.e. brain size) of 1,800 ml that far exceeds the average value of 1,400 ml seen in humans today. But despite this "modern" brain size, other aspects of the cranial vault such as its low, sloping profile make it look much more like Homo erectus, which lived in China as far back as 1.6 million year b.p. Even more intriguing is the back of Xuchang 1’s skull, which has a modest torus with a depression above it, known as the suprainiac fossa. This trait is not known in Homo erectus or Homo sapiens, but is very common among Neanderthals from West Asia and Western Europe.
The Xuchang fossils have anatomical features that do not look like they are derived from African Pleistocene populations, nor do they always look like modern-day Asians. Anatomically, they are a combination of Asian Homo erectus, Western European Neanderthals, and early Homo sapiens. While this specific combination of traits is not widespread, the overall pattern of morphology that is a mosaic of Eastern and Western Eurasian as well as African is seen in other Chinese fossils from the Middle and late Pleistocene such as Dali, Maba and Xiujiayao. This suggests that the evolutionary forces shaping this morphology, including gene flow and genetic drift, were acting throughout China.
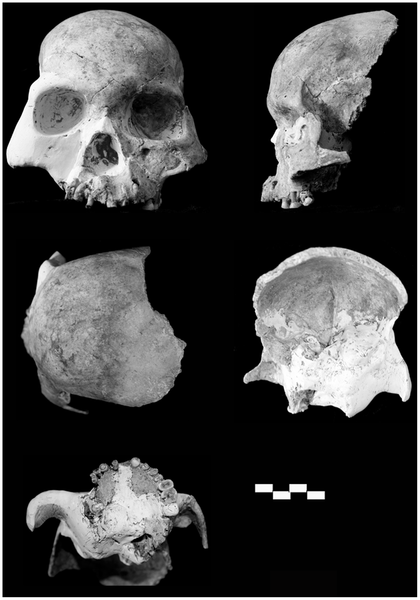
Tianyuandong and early Homo sapiens diet and DNA
The Tianyuandong specimen (known as Tianyuan 1) from the Tianyuan cave in Zhoukoudian village, just six kilometers from the famous Homo erectus site, is the earliest directly dated Homo sapiens from China . Enough collagen was preserved in its bones to date it to between 42-39,000 years b.p., which is roughly the age of the youngest Upper Cave specimens at the nearby original Zhoukoudian site. The preserved collagen also allowed researchers to extract ancient DNA from Tianyuandong, and even to study its diet.
Anatomically, Tianyuan 1 is clearly Homo sapiens despite the fact that its age coincides with the existence of other populations that are believed to be separate species such as Neanderthals. For example, it is not particularly robust aside from having relatively large front teeth (which is common among non-agricultural populations because they usually have tougher diets). The diet of Tianyuan 1 was reconstructed through studying the chemical components present in its teeth. Anthropologists look at the isotope values of bone collagen because these are related to the isotope values present in the food that a person consumes during their lifetime. For example, a higher proportion of heavier vs. lighter isotopes of carbon (13C/12C) indicates that the main source of dietary protein was marine vs. terrestrial plants and animals. In the case of Tianyuan 1, the carbon (13C/12C), nitrogen (12N/14N), and sulfur (34S/32S) isotope ratios all point to freshwater fish as a main component in the individual’s protein diet. This tells us that as far back as 40,000 years ago, humans knew how to fish and exploit coastal resources for their survival.
The Tianyuan 1 specimen is the only Pleistocene Chinese fossil to have its ancient DNA analyzed thus far. Its genomic sequences were compared to 11 individuals from today, which were used to represent populations from Africa, Europe, Asia, and South America . This sample size may sound small, but when dealing with genomes it is reasonable to assume that one individual possesses the common alleles (i.e. gene variants) for that gene pool. The Tianyuan genome was more similar to the two Asian individuals from the present-day than it was to the European, African, or South American. So even though Tianyuan 1 is from a 40,000-year-old population, it isn’t genetically distinct from people living in China today. Put another way, there has been regional continuity between the genomes of ancient and present-day eastern Chinese.
The Xuchang and Tianyuan sites are important for understanding how the traits we see in Asia came to be established. Between the 100,000-year-old Xuchang and the 40,000-year-old Tianyuan fossils, a great deal of population change was happening throughout Eurasia, Africa and Australia. At 100,000 years b.p. Homo sapiens are established in China (Zhirendong), West Asia (Israel), and in Northeastern (Ethiopia), Northwestern (Morocco), and Southern Africa (South Africa). Archaeological evidence suggests they arrived in Australia by 65,000 years b.p. . But in Western Europe, Neanderthals dominate the landscape throughout this time period, until as recently as 35,000 years b.p.. And although by 250,000 years ago, Homo erectus is no longer seen in China, the name for the fairly rich record in China of transitional erectus/sapiens-like fossils dated between 250,000 years b.p. and the earliest known Homo sapiens at 100,000 years b.p. is still unclear . Some refer to them as Homo heidelbergensis and see them as a separate branch on our evolutionary tree. Based on my work in India, China and Indonesia I think they are part of a continuous lineage in Asia from the Middle to the Late Pleistocene, and possibly even longer.
Braided streams: A new model to explain human evolution
To better clarify the evolutionary processes that shaped these populations beyond the simplistic binary of replacement vs. continuity, my colleague Wu Xinzhi at the Institute of Vertebrate Paleontology and Paleoanthropology in Beijing developed a model in the late 1990s that seems to have a lot of explanatory power for China. It is known as "Continuity with Hybridization" . Wu was trying to make sense of the mosaic of traits he observed in the Middle Pleistocene fossils in China, many of which he excavated and was the first to analyze. Based on comparative anatomy, he saw that in some respects they had features that were common in European but not African fossils. In other respects, he saw clear connections with earlier Chinese Homo erectus. Perhaps most importantly, he recognized that while there was a lot of variation among the Chinese fossils, they were still unified by a handful of features.
This prompted him to propose a model of gene flow through time that he described as a braided stream. Populations in what is present-day China would experience periods of "isolation" that would lead them to be shaped by very specific local forces, much the way streams are shaped by local landscape and climate. Over time, these lineages would come back into contact with other populations, exchanging genetic material for a period, which would create commonalities among them. This process, Wu argues, continued throughout the Pleistocene and shaped human evolution in China.

The ideas that Wu raises in this model are intriguing. To understand it, we have to start thinking of human populations along a continuum. At one end of the spectrum are local communities where individuals choose mates. At the other end are entire regions of people who share a broadly defined ecological zone, such as all individuals living in eastern China north of the Han River. At the very local level you may have individual groups occasionally become isolated and experience minimal gene flow from the outside, perhaps even go extinct. This can happen when population densities are low. In this model, those localities represent individual "streams" within a larger braided river. They are following their own evolutionary trajectory for a time and are not subject to the forces shaping the vaster regional population – in this case, all of present-day China. Eventually, these local groups are reconnected with the larger gene pool (the trunk of the river) and exchange genes again. As a result, we would see some unique traits develop at a local level during those periods of isolation, but would also see the persistence of broader regional trends through time. In other words, there would be continuity with hybridization – with hybridization referring here to gene flow at the larger regional level, between western and eastern Eurasia or Africa. This model contains elements of existing ones but was developed independently using data just from China. So it is not meant as a refutation of the debate that has dominated western science, nor to explain all of Homo sapiens evolution, but rather to apply regional expertise and knowledge to understanding evolutionary processes in China. Whether it is applicable to other regions can be tested, and it has been proposed for other areas as well, but the one-size-fits-all era of hominin evolution models is over.
Maludong and Longlin: Southwest China at the end of the last glacial period
Two sites where we can apply the predictions of the Continuity with Hybridization model are the Maludong and Longlin caves in Southwest China, which have been known for the past 20-30 years. A partial cranium was found in Longlin in 1979 by a miner, and a number of different skeletal elements were excavated from Maludong in the 1980s and again in the past five years . Like the Xuchang crania, the Longlin and Maludong fossils have a suite of traits that are not seen in other fossil populations although individually each trait is present in Neanderthals, Homo erectus, or early African Homo sapiens. These two sites are much younger than Xuchang – they date to around 11,000-14,500 years b.p.. This period is known as the Terminal Pleistocene because climatically, it spans the end of the last glacial period and the onset of the warm period we live in now, known as the Holocene. In China, the climate in the region went through a cool and dry spell, and communities were just on the cusp of developing a growing dependence on plant foods that they would begin to domesticate by around 10,000 years b.p.. This would mark the beginning of agriculture in the region.
In this context, the partial human skull from Longlin Cave and the fragmentary skull, teeth and jaw bone from Maludong shouldn’t necessarily be expected to look like other fossil populations that lived tens if not hundreds of thousands of years before. What makes them remarkable is that they do not align with any living East Asian populations, or in fact with any Homo sapiens populations at all from the Later Pleistocene. They have brain sizes that are average for our species, but their skulls are overall fairly narrow in the front, and their cheek and teeth regions are large and flared. Even the teeth themselves are fairly robust. This may be related to diet. Maludong means "red deer cave", named for the large amount of red deer bones that were found along with the human remains. Presumably this population relied on game as a major food source, which is in contrast to older populations such as Tianyuandong, which relied upon freshwater fish.
Because of the number of archaic traits on the Longling and Maludong crania, the researchers who analyzed them argued that they were late-surviving archaic hominins, or they were the product of multiple waves of "colonization" into East Asia by Africans exhibiting evidence of hybridization between "archaic" and "modern" humans. Both of these scenarios assume that the populations of Southwest China during the Late Pleistocene were a separate species from Africans, and base this assumption on their appearance – specifically on the fact that they do not have high foreheads and lightly-built faces, traits that were first defined as "modern" based on (present-day) European morphology and then identified in fossils from Northeast Africa and West Asia.
Additionally, the definition of species is subjective, with some biologists using the biological species concept and other using the evolutionary species concept or any other number of concepts . An alternative interpretation of the Longling and Maludong fossil morphology proposed by Rosenberg and Wu gives less weight to the archaic traits and points out that they possess a mosaic of features, each of which is also seen in other Eurasian populations (much like the Xuchang 1 specimen described above) . Viewed through the lens of the Continuity with Hybridization model, these populations are not descended from a very recent presence of Africans coming into Asia, nor are they part of an uninterrupted continuous lineage confined to China over the past 1.6 million years. Rather, they are the product of a number of evolutionary processes that included gene flow between eastern and western Eurasia, as well as southeast and north Asia.
By focusing on evolutionary processes instead of labels, we can get a better sense of the truly interesting questions about the history of our species. We don’t just want to know what things looked like so that we can categorize them, we want to know what forces shaped the traits that we are seeing. There were many paths that different populations took to get to where they are today, and our goal is to understand how much gene flow, genetic drift, and natural selection shaped us. This means moving beyond shoehorning all of this information into 30-year-old models of “Out of Africa” and “Multiregional Evolution.”
What happend to Homo erectus in East Asia?
When the models of "modern human" origins were first developed in the 1980s one driving issue was: did Homo erectus in Asia go extinct, or evolve into Homo sapiens? The success of the Out of Africa model meant the default assumption was that Asian Homo erectus was an evolutionary dead end. However as all of these sites show – particularly Xuchang and Tianyuandong – gene flow throughout Eurasia was instrumental in shaping early Chinese Homo sapiens, but so too was evolutionary continuity within the region throughout the Pleistocene. This implicitly includes the time period of Homo erectus.
Paleoanthropologists in China have been aware of the continuity between Homo erectus and Homo sapiens without wholesale replacement for decades . With recent advances in dating techniques and ancient DNA analyses, it is clear that humans have been living there for the past 1.6 million years and that they were biologically highly variable, as would be expected for a land mass that is 9 million square km large. It is also clear that they were not isolated. The fossil evidence from the Middle Pleistocene showing a mix of Homo erectus and Homo sapiens traits, along with the complex morphology in the fossils discussed here that have affinities with Neanderthals, early Homo sapiens, and living Chinese, all solidly refute the idea of Asian Homo erectus going extinct and being replaced by Homo sapiens. One can story this as Homo erectus populations becoming assimilated in the course of millennia by Homo sapiens groups (i.e. they contributed to the ancestry of Homo sapiens in the region), much like the Neanderthal populations in western Eurasia, or of Homo sapiens having an incredibly deep and gradual evolutionary history in East Asia that cannot be pinpointed to single origin or time.
Regardless of how one views the boundaries of past species, it is clear that populations in eastern Eurasia throughout the Pleistocene were shaped by gene flow, random genetic drift and natural selection – the basic forces of evolutionary change. Just how much of each of these forces was at play remains to be understood. Gene flow acts to keep two populations from differentiating, while genetic drift tends to enhance differences between groups. At various periods, genetic drift in an isolated population (a braid in the stream) would have allowed for unique features to develop while at other times, gene flow (the joining of two braids in a stream) would have worked to assimilate various Eurasian populations.
With each new discovery and the application of new technologies, we are finding out about the evolutionary history of our species beyond Africa. This is the kind of holistic picture we need to truly understand who we are and where we came from. The story will be, like us, very varied.
Want to know more?
- Athreya S and Ackermann RR (2019) Colonialism and Narratives of Human Origin in Asia and Africa. In Porr M and Matthews J (Eds.) Interrogating Human Origins: Decolonisation and the Deep Past Routledge
- Rosenberg KR and Wu X (2013) A river runs through it: modern human origins in East Asia. In Smith FH and Ahern JC (Eds.) The Origins of Modern Humans: Biology Reconsidered John Wiley & Sons
Other Articles by This Author
Further Articles in Paleobiology
 This work is distributed under a Creative Commons Attribution ShareAlike 4.0 International License (CC BY-SA 4.0). Its use, distribution and reproduction in other forums is permitted provided both the original author(s) and Capeia are accredited, and the license is unchanged (CC BY-SA 4.0). Images or other third party material in this article are covered under the article’s Creative Commons license unless otherwise indicated; if the material is not covered under a Creative Commons license, users will need to obtain permission from the license holder before it can be reproduced.
This work is distributed under a Creative Commons Attribution ShareAlike 4.0 International License (CC BY-SA 4.0). Its use, distribution and reproduction in other forums is permitted provided both the original author(s) and Capeia are accredited, and the license is unchanged (CC BY-SA 4.0). Images or other third party material in this article are covered under the article’s Creative Commons license unless otherwise indicated; if the material is not covered under a Creative Commons license, users will need to obtain permission from the license holder before it can be reproduced.
Responses